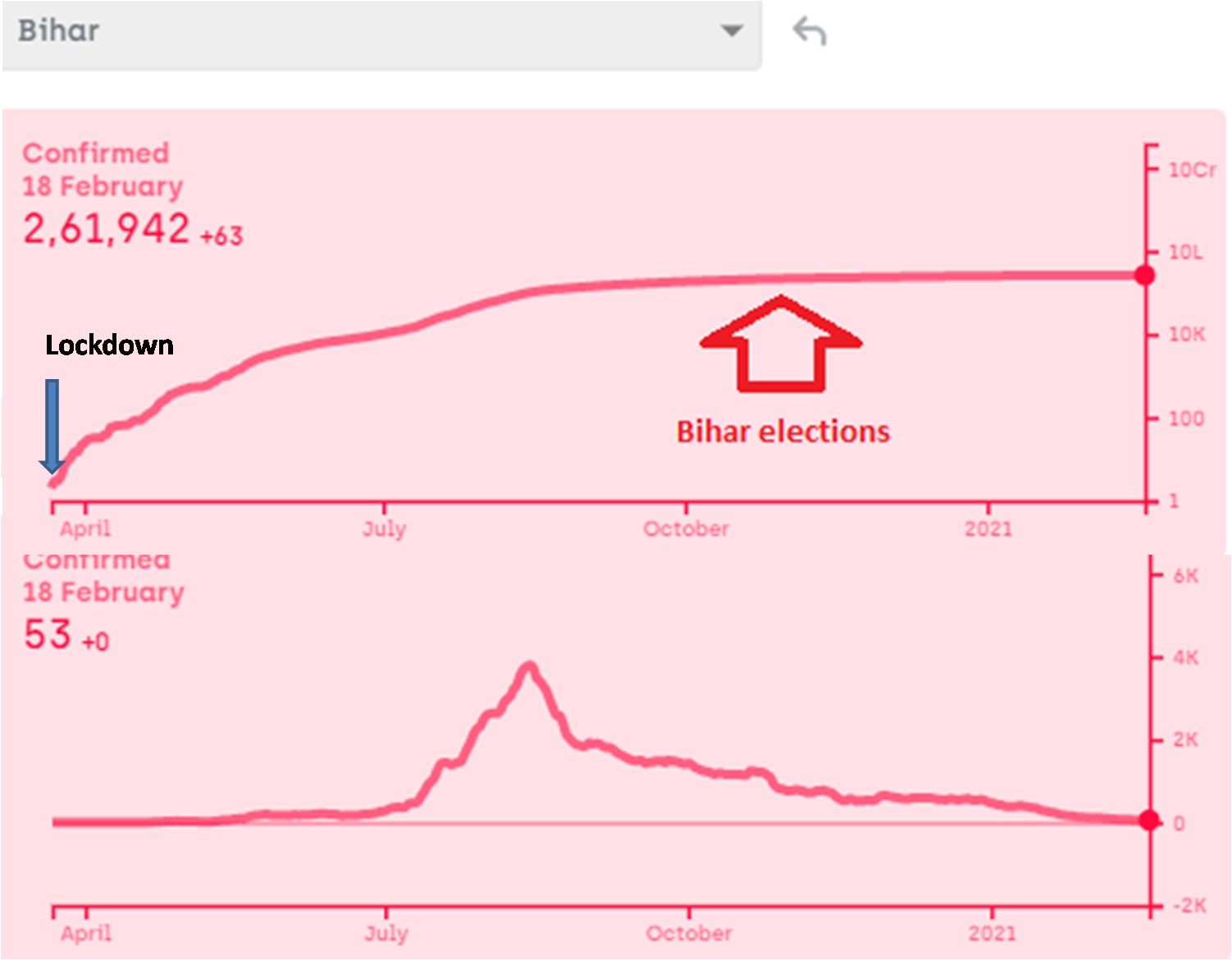
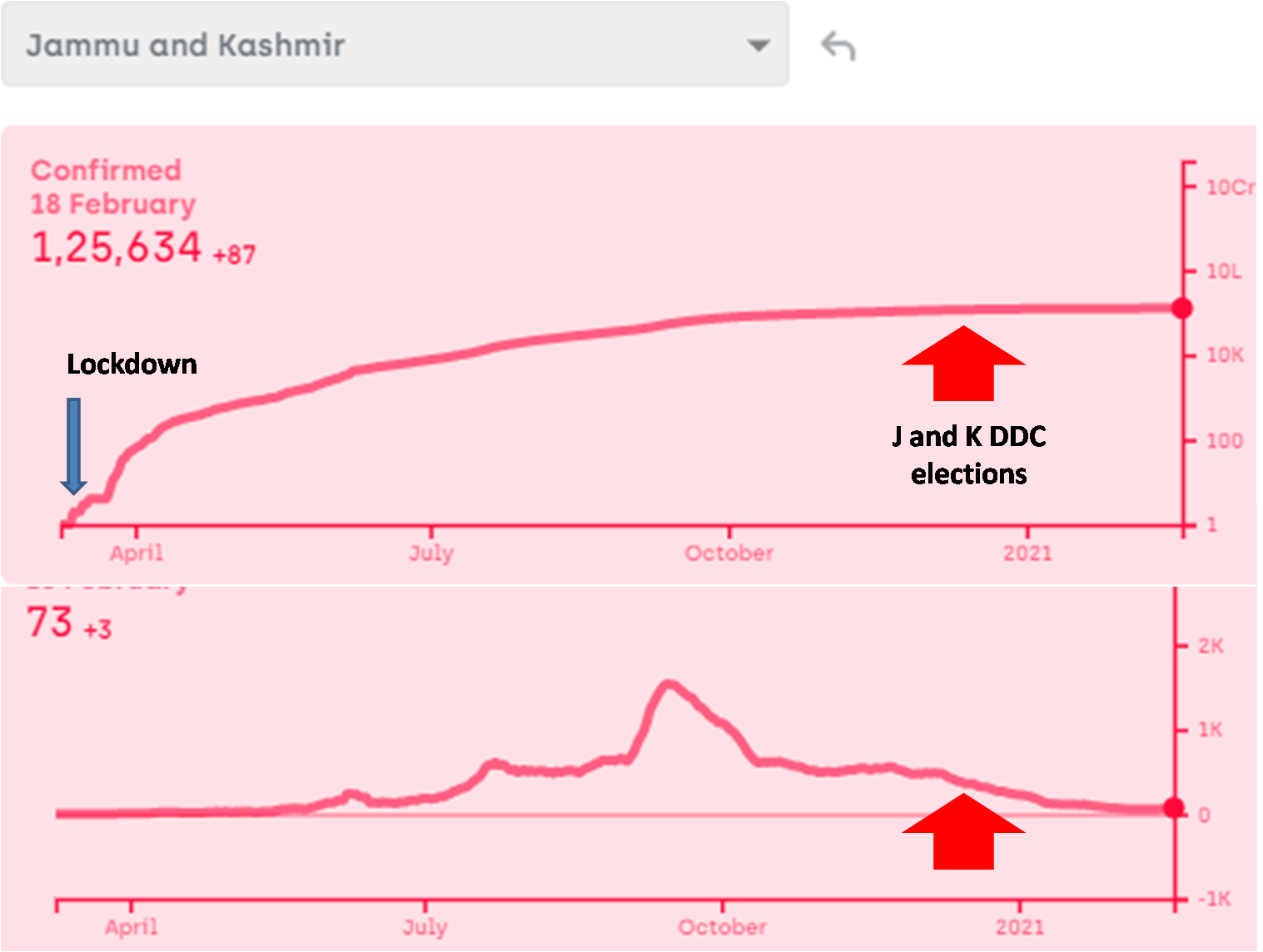
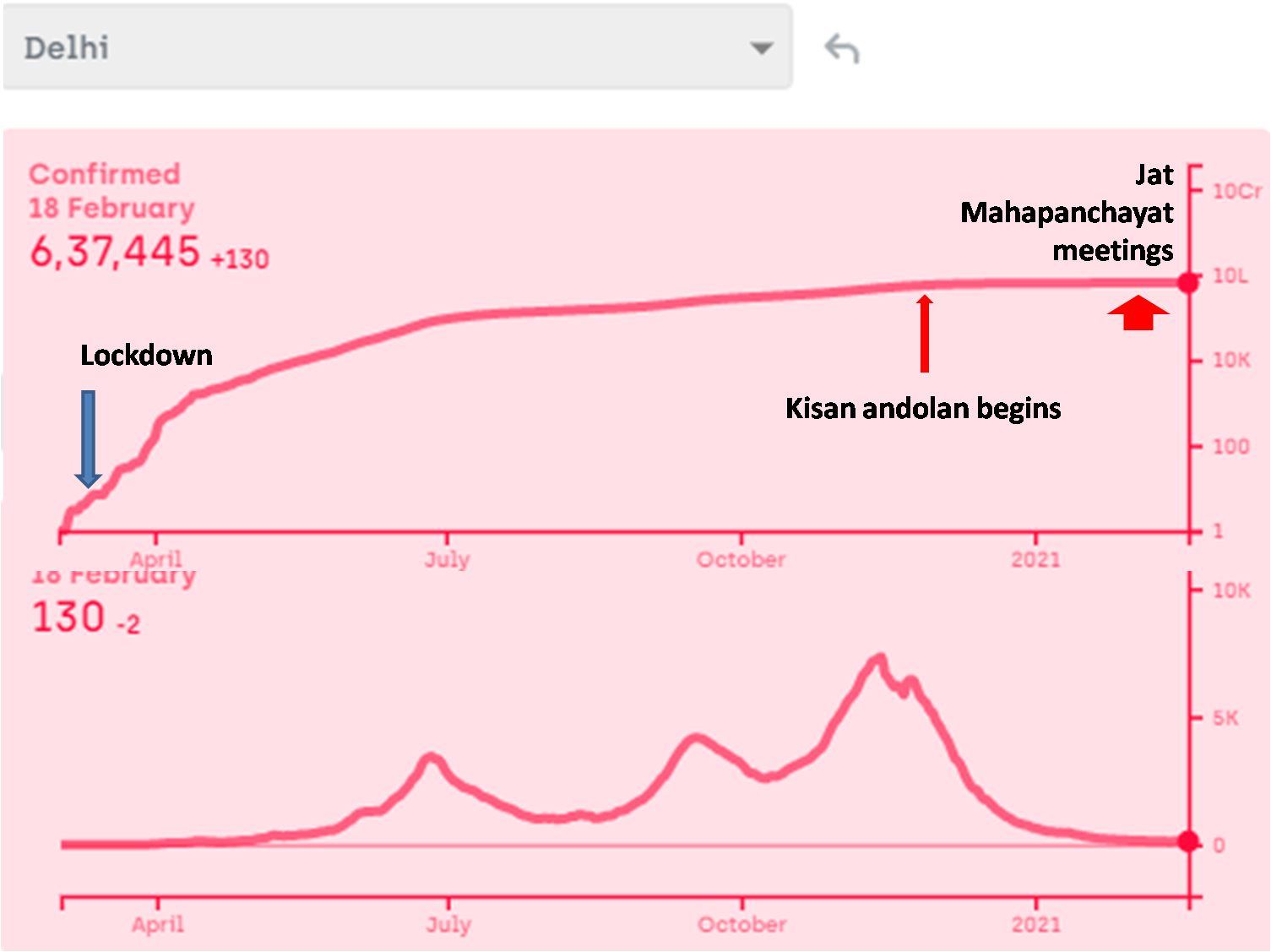
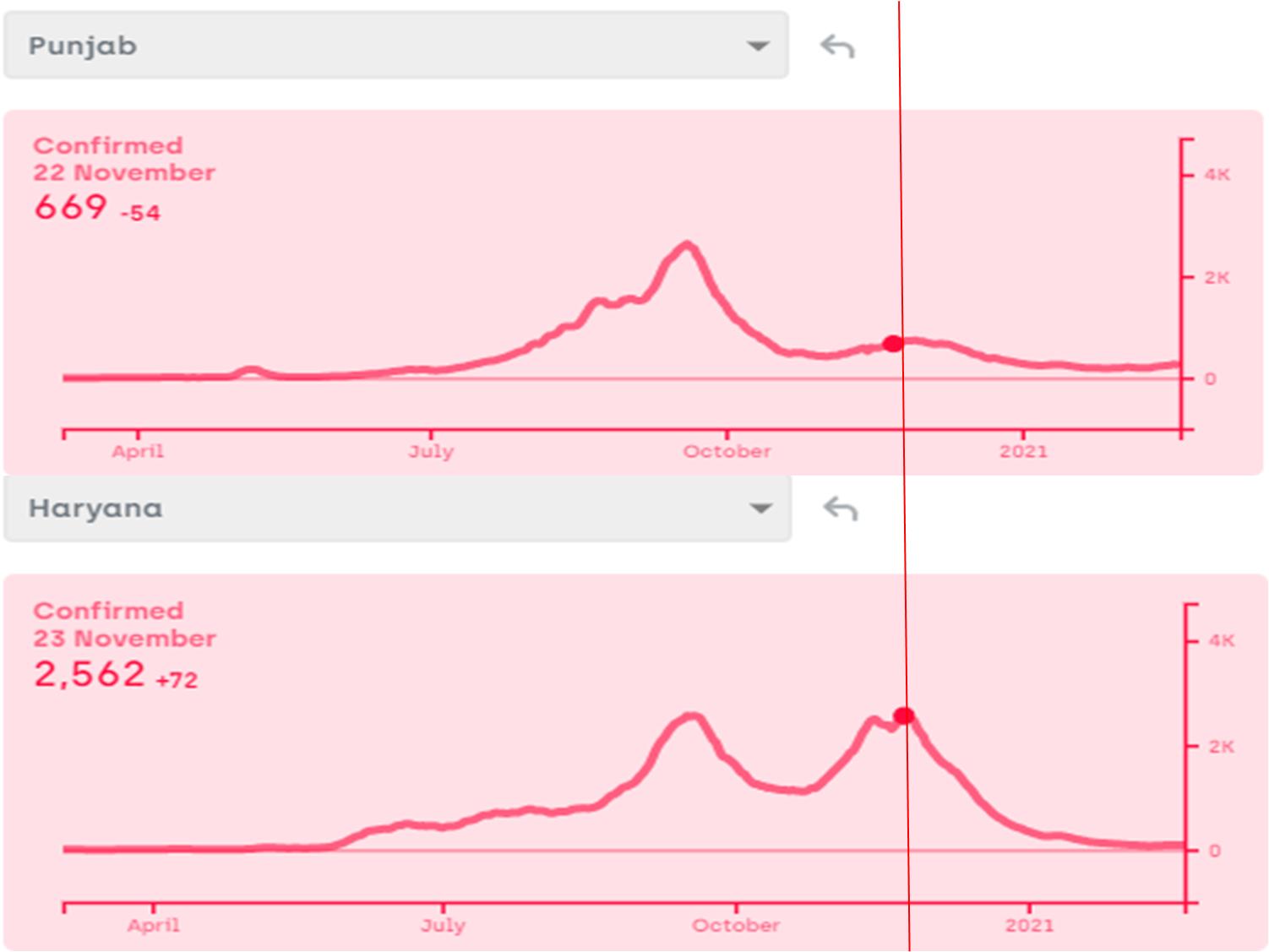
I am fully aware that nobody would listen to me and change anything at the policy level, but as a science minded citizen I am going to write here what I feel is the most logical and effective plan to deal with a third, fourth or subsequent waves of the pandemic. In some countries, a third (or even a forth) wave has certainly appeared already, in spite of vaccination, but the death rate is substantially lower. Since we have invested a lot in vaccines, it would please us to say that the death rate came down because of vaccination. This might be partly true, but it has been coming down even prior to vaccination, although not without hick ups. Interpreting these hick ups will not only be interesting but important for handling future waves wherever and whenever they appear. My views of successful handling of the third wave depend upon my interpretation of the patterns so far. Therefore I am putting down both simultaneously here.
I have an eight point program to effectively manage the pandemic hence forth, including any surge, if, when and wherever it occurs.
- Give up the Don Quixotic attempt to “break the chain”: We have been often hearing phrases such as “break the chain”, without understanding what it means. How are we supposed to break the chain, i.e. interrupt one round of infection cycle? By preventing the transmission from currently infected individuals to susceptible individuals. If we can do this for one complete cycle, i.e. about 3-4 weeks, the epidemic will stop once for all. That is the true meaning of “break the chain”. Nobody has successfully broken the chain anywhere in the world so far except where there were only a handful of cases at that time. Such a situation does not exist anymore in any country with only half a dozen possible exceptions. According to a WHO report (https://apps.who.int/iris/handle/10665/329439), none of the measures suggested including social distancing, masks, sanitizers, lockdowns, school closure, work place restrictions, entry exit screening individually or collectively have evidently ever stopped transmission. They can at the most reduce transmission rate by a few percent. Even the flawed reports that claimed these measures to be effective in the first phase of Covid did not show efficiency high enough to break the chain. So the call to “break the chain” was Don Quixotic right from day one. But it was promoted as a rhetoric and people believed it with religious faith.
This reminds me of a story. A spiritual healer told people, if you do such and such ritual with full faith, the disease will vanish. Saying such a thing is very safe for the healer. If the disease really vanishes, he can take the credit. If it doesn’t, he can say you didn’t do it with full faith, therefore it didn’t work. The same applies to breaking the chain by social distancing. If by chance, the incidence goes down, it is because of that. If it doesn’t, it is because people did not follow the rules properly. Therefore the health authority are safe in advocating it. Going by such a logic the concept that preventive restrictions can arrest transmission and break the chain can never be proved wrong. It has no falsifiability and therefore by Karl Popper’s definition is not science at all.
In order to arrest transmission, we should first know how the virus is transmitted. Even after one and half years there is no agreement on the relative importance of droplets, aerosols and surface transmission. What is a safe distance has never been determined. Some studies say two meters some say ten meters. So even theoretically what measures can be expected to prevent transmission is uncertain.
Instead of trying to arrest transmission of respiratory virus, which is hardly in our control as we were made to believe, we need to focus on something that is in our hand. Good patient care and vaccination is certainly in our hand. Both are not magic solutions but they can certainly save many deaths and that is evident in data.
In the history of infectious diseases, out of the several dozen viral diseases caused by hundreds of variants, only small pox has been successfully eradicated and polio might have been. Human efforts have not been able to get rid of any other virus so far. But we are not afraid of most other viruses anymore, because we managed them successfully in some other way. They are still there but they no more kill as many people. That is the most likely fate of SARS-Cov-2 as well. How to achieve this as fast as possible should be the prime R & D and health policy question. But such a question is hardly being addressed anywhere. If we are still talking about breaking the chain, getting rid of the virus, keeping it away forever or eradicating it, we are the reincarnation of Don Quixote.
2. Change the testing strategy: tests to detect active infection serves three possible purposes (i) help in deciding the course of treatment (ii) contact tracing and isolation of potential spreaders for arresting transmission (iii) give useful epidemiological data. A careful look of patterns over the last 15 months through most of the world shows that testing, as it is being done, has not served any of the three purposes. The course of treatment of a patient is mainly decided by the symptoms. Being or not being Covid positive has little role in deciding the course of treatment most of the times. If, in a rare case, the treating physician thinks he needs to know the test report to decide the treatment, a test facility should be available. But to do millions of tests is unnecessary to serve any clinical purpose.
Contact tracing can be effective only when there is a small number of cases and only a small proportion of asymptomatic and undetected cases. Above a threshold incidence, it simply stops working (https://milindwatve.in/2020/07/20/covid-in-india-why-the-strategies-need-to-change/). At this stage of the pandemic with a large number of undetected cases, contact tracing to prevent transmission is equally Quixotic. So continuing this effort is not worth.
On the third purpose, the way the tests were being done so far, the data generated has been highly biased and quite unreliable for any useful epidemiological purpose. So tests have not served any of the three intended purposes.
So I would suggest the following strategy for tests. (a) For clinical purposes, only when a test is likely to influence the course of treatment, as decided by the physician it should be done. (b) For generating good epidemiological data, instead of doing millions of tests haphazardly, let a cluster of institutes be given a mandate and adequate funding to do well-designed sample surveys which will generate useful epidemiological data. What is important for epidemiology is not the total number of cases, (which nobody is likely to know anyway, all numbers given so far are wrong!!) but faithful representation of the trends. This needs well thought out sampling design, not simply more massive testing. Attempt to detect every infected person by a test is also a quixotic effort and needs to be given up as fast as possible. A well designed sampling strategy will be immensely more useful, simultaneously saving cost as well as panic and confusion in the minds of people. Better data will be available to scientists and all the data mismanagement can be eliminated.
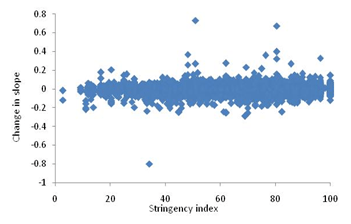
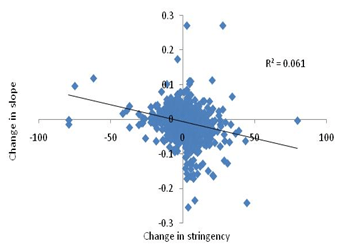
3. Lock the lockdowns: Detailed and careful global analysis gives little evidence that lockdowns help reducing the transmission of the virus substantially, making them worth paying all the economic, educational and social costs. The distribution of the costs over the society is highly unequal. The decision makers themselves pay hardly any cost. Focused protection of the vulnerable groups and normalizing all other life needs to be the priority. At the most large avoidable gatherings can be restricted. So far there is no convincing and consistent evidence that such gatherings have increased the transmission of the virus, but one may do that if the events are not perceived essential.
Travel restrictions have led to another strange problem. As we reduce substantially the transmission across countries and regions, the epidemic curves take different shapes in different regions. When the prevalence declines in one area, it is on the rise in another. These non-synchronous surges ensure better long term survival of the virus. So it is quite likely that travel restrictions actually help the virus more than us. Also the virus will evolve differently in different parts of the world adding to the uncertainty of the course of the pandemic.
4. Invest in people oriented patient care practices: A good practice to face a calamity is to hope for the best but simultaneously prepare for the worst. Going by the worst of all mathematical predictions, it is necessary to make patient care facility available at every corner of the world. The lockdowns did two kinds of damages to prevent this. One is to give a false assurance that masks and social distancing will prevent the spread, so less attention was given to development of patient care facilities. The second was that the lockdowns made the economy collapse in some areas so badly that investment in the facility and availing the facility also became difficult.
Furthermore, at least some of the patient care practices of the first and second wave were counter-productive. Over the last week I have been traveling in some tribal interiors and talking to people about their Covid experience. My impression is that the wave swept across even the remote areas and almost everyone was sick during the peak period. But people were reluctant to get tested and admitted to hospitals for multiple reasons. One is that the government hospitals were full and private ones were unaffordable. After losing jobs in the lockdown, even the cheapest of the hospitals and transport to the hospitals was not affordable to many. Other, even more important was our patient quarantine practice. Quite often a person admitted to a Covid ward was not seen by its relatives again, not even the dead body. Culturally and emotionally, death rituals are extremely important. Denying people even a sight of their dear ones even after death, led to increased reluctance to admitting and even testing for Covid. Seriously infected patients remained at home spreading the infection further. The secretive funerals may have been intended as an extra precaution to prevent spread, but in reality it might have been counterproductive. This illustrates the need to understand people and make the health care practices people oriented.
5. Directing evolution of the virus: It is very clear that the virus is evolving very rapidly. We have lots of data on mutations but almost no insights into what these mutations mean clinically. But we see simultaneously that death rates have decreased although apparent infectivity may have gone up. There is a set of conditions under which as the virus evolves in a host population its virulence goes down although infectivity may go up. In the context of Covid, two conditions are crucial for evolutionary changes in virulence. (i) If virulence is related to viral replication and transmission, the virus has a selective advantage in being virulent. But in Covid 19, the correlation between disease severity and respiratory viral load is very poor as shown by multiple studies (DOI 10.1016/s2213-2600(20)30354-4; DOI 10.1101/2020.07.20.20157792; https://jamanetwork.com/journals/jamainternalmedicine/fullarticle/2769235). This means that there is only a marginal advantage to the virus in being more virulent. (ii) On this background, if the serious cases are effectively quarantined and the mild and asymptomatic cases remain intermingled with the population, the milder variants will have better chance to spread. This will eventually lead to loss of virulence. The rate of this loss will depend upon how we handle the epidemic. (a) If we attempt efficient contact tracing so that all cases including asymptomatic ones are always diagnosed and detected, we are not giving any selective advantage to the milder forms. In such a case a more virulent virus will evolve because there is at least a weak correlation of virulence with viral replication. (b) If we allow the mild and asymptomatic cases to mix with the population, but effectively quarantine the serious cases, we give a selective advantage to the milder variants. (c) If, there is no effective isolation at all, then again higher virulence is more likely to evolve. By carefully managing and differentiating between cases we can boost evolution of the virus towards loss of virulence. This might be bad in the short run but good in the long run and I suspect, the only strategy that has high chances of effectively ending the epidemic.
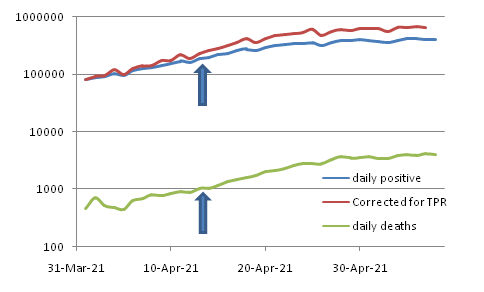
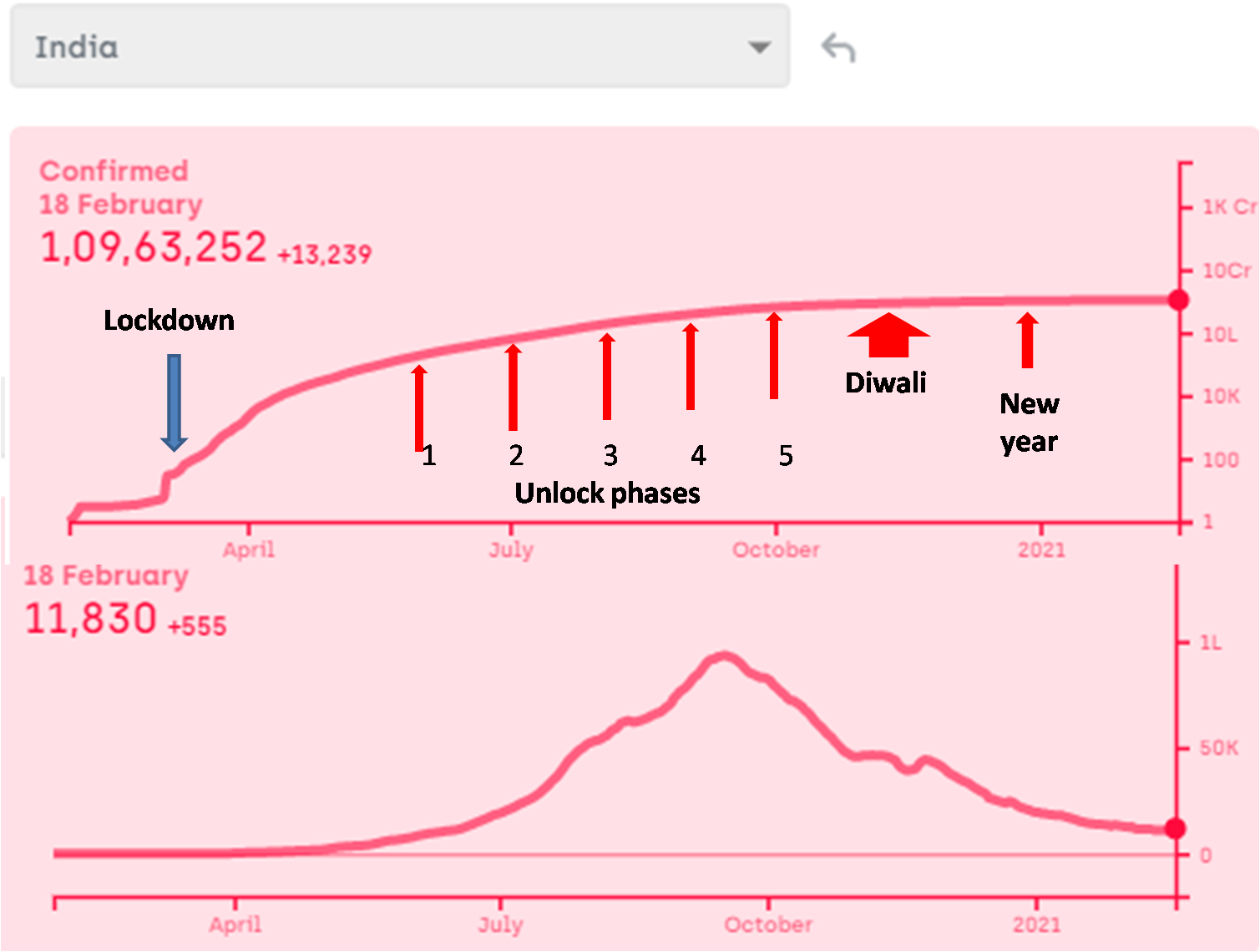
Interestingly nobody in the mainstream even talks about evolution of virulence. On a few occasions responsible people talked about it and they were immediately trolled so heavily that evolution of virulence became a taboo in this field. But death rates reflected in different indices have been coming down, with some transient trend reversals. The trend reversals also can be interpreted logically. In India, the death rate at the beginning of the second wave was only one fifth of that in the first wave. But it gradually increased as the second wave peaked. I can see two possible reasons for it. One is the saturation of hospitals and patient care facility which clearly increased the death rate. The other is more subtle. As explained above, in remote rural India, the funerals under isolation, often without even informing relatives, led to increased reluctance to admitting and even testing for Covid. Many serious cases remained and died at home. In the absence of isolation of severely infected individuals, the virulent strains must have had a good chance of spread. Thus cremation under strict isolation may be intended to prevent transmission, but it actually ended up increasing transmission, that too of the more virulent forms.
Strengthening host immunity also affects the evolution of the virus in subtle ways. Immune response is costly for the host physiology. The immune response has evolved to see that the cost of immune response should not be greater than the cost of invasion itself. Therefore the body does not give an all out immune response to all pathogens all the time. The investment in an immune response is carefully weighed against the intensity of invasion. A virulent virus will evoke an all out immune response and thereby destroy itself in an immune-competent host. A mild virus is more likely to escape intense immune response and thereby survive better. So as the average immunity of the population goes up, the virus evolves to become progressively milder. Vaccination drive serves this purpose the best, rather than the proposed quixotic eradication of the disease, or preventing infection or preventing subsequent waves which it hasn’t. Signs of third or fourth wave are clearly seen in countries with good vaccination coverage as well. Directing viral evolution is going to be the truly useful role of vaccination. Otherwise we have seen that the vaccines neither prevent subsequent waves nor stop new variants from arising. In fact greater the proportion of vaccinated, greater the selective force for new variants.
Given the small genome and huge global population of the virus, the evolution of the virus is most unlikely to be mutation limited. It is expected to be selection limited and therefore it is possible to manage selective pressures on the virus so that it rapidly becomes milder and remains just another coronavirus.
6. Fitness drive: Global data and multiple published studies have shown that physical and physiological fitness, independent of age, has been keeping serious symptoms and death away. There is consistent global data on fitness relating to Covid survival (see for example, https://bjsm.bmj.com/content/early/2021/04/07/bjsports-2021-104080). Sedentary lifestyle is a high risk factor. During the first and second waves we forced the gyms, playgrounds, jogging parks and swimming pools to shut down. This is very likely to have increased the susceptibility of the population. Promoting rather than closing down fitness activities would be the right strategy if there is yet another wave. Fitness is the most reliable way of surviving the infection. Closing down fitness activities was the biggest mistake that should not be repeated in any case.
7. Make up for the loss in education? There is no need for school and college closure. That is the age group least affected by the virus. There is little evidence that closures helped. But even if we reopen now, the hardest thing to do is to recover the educational loss already incurred. Recovering from the economic loss might be easier than recovering from the educational loss of an entire generation. How to recover is one of the toughest questions and needs a lot of thinking to go in. Perhaps there may not be a single solution to all levels, all schools and all subjects since details matter a lot. But certainly a focused special drive is needed for making up the lapses in education.
8. Research questions to focus on: The pandemic has exposed the inadequacies of the way research community works. We have failed to identify and address the most important questions, recognized alternative possibilities, making specific local context based policies, cross question and recheck earlier conclusions, challenge rhetoric and keep on improving our understanding of the disease. While we did see some good science during this time, many important questions were almost completely left out. The precise mode of transmission of the virus is still not clear including the importance and dynamics of aerosols versus other modes of transmission and their context specificity. Realistic evaluation of the performance of various preventive restrictions in the context of different localities has been seriously lacking. There were some early attempts that were seriously flawed. Later there seems to be nothing published. It is possible that public health policies that work in the short run may fail to work or even become counterproductive in the long run (see for example, https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1008292). But we have been emphasizing on the same preventive measures, for the success of which we have no reliable evidence. There is need to reassess the efficiency of all the measures periodically, rethink and redesign the policies accordingly. Substantial amount of research needs to go in such an effort in every country because the success will depend upon local conditions which are different in different countries/regions/cities. What works in one city may not work the same way in another. Assuming what worked in New Zealand will work in India is being superstitious.
The epidemiological models have largely proved themselves inadequate, to put it mildly. The reason is that they are oversimplifying the population processes. For example almost all models treat immunity as a binary variable. An individual is either susceptible or immune, which is far removed from reality. This binary perception of immunity has led the illusionary herd immunity concept. Cross immunity offered by common cold and other milder viruses, immunity by repeated subclinical exposure to the virus are not considered at all by the models. These can potentially be factors chat change the course of the epidemic even qualitatively. Building such models need not be too tough, but needs an appreciation that current models are failing and we need to incorporate at least some of the complexities of the immune mechanisms in the model by making immunity a continuous and perhaps multidimensional variable. I haven’t come across attempts to build such a model. Not because it is difficult but because the vision is surprisingly absent.
Most important, we need a lot of realistic research on human behavior. The success of any public health policy depends upon whether it is behaviorally sound, how people would interpret it, what will be the consequent perceptional and behavioral changes in the short and in the long run, whether people will follow the advice, will it become a ritual and lose its original meaning and so on. Study of human behavior needs to be an intrinsic part of policy making in every field. For public health it makes lot more sense.
Boosting research in the right direction is also a complex behavioral problem and needs to be perceived that way. This is a more fundamental question of research culture itself about which I have written many times and will keep on writing. So I won’t expand on it here. But I would certainly hold the research community ultimately responsible, if the governments are following any strategy without sufficient evidence base, without repeatedly getting evaluated and in turn causing more harm to people than the disease itself.